Back to article: A game of substrates: replication fork remodeling and its roles in genome stability and chemo-resistance
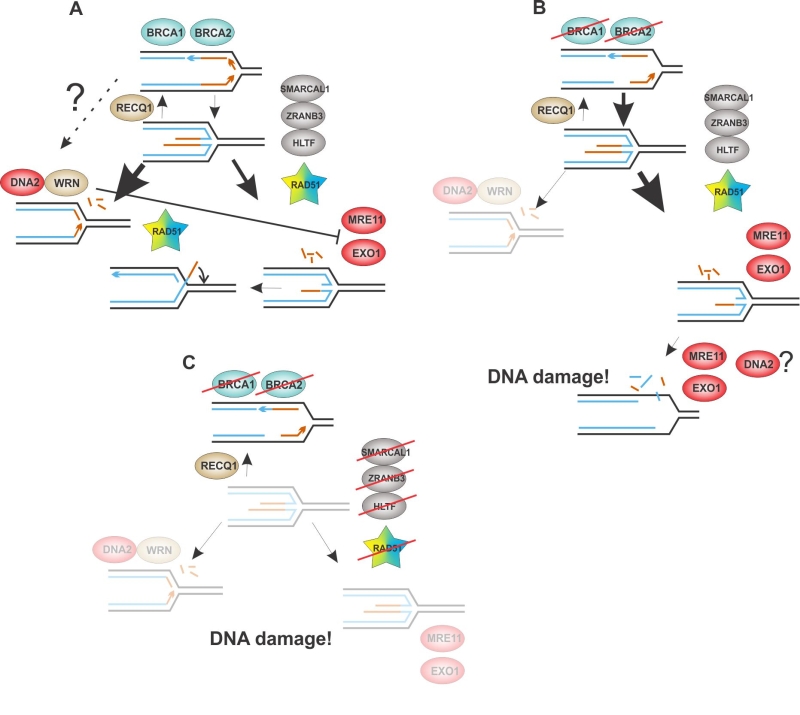
FIGURE 4: A model for two interlinked branches of fork resection and their alterations upon mutational loss of the involved proteins. In a wild type cell the MRE11, EXO1 branch may be less prevalent than the DNA2/WRN branch, depending on the cell background. An inhibitory feedback from WRN to MRE11 may contribute to this arrangement. Fork resection may be limited by restorative branch migration of the fork junction, or reannealing of the 3’ overhang in a D-loop-like strand exchange (A). In BRCA1 or BRCA2-defective cells, the MRE11, EXO1 branch dominates and WRN is no longer a contributor. Fork resection proceeds beyond the four-way junction, either directly, or via cycles of reversal/resection, leading to DNA breakage either directly or via segregation of under-replicated DNA (B). Inhibition of fork reversal suppresses fork resection but also decreases stalled fork stability and ability to resume replication. DNA damage may result from fork collapse and segregation of under-replicated DNA and is exacerbated by BRCA1 or BRCA2 deficiency (C). Faded areas in the cartoon indicate pathway inactivity. See text of the sections “RAD51 versus the nucleases: reversal, protection, resection?” and “The nuclease crosstalk and the curious case of WRN” for a detailed discussion.